TEMA BROJA
Dr Gordana Jovanović
Živi svet praistorije / Evolucija veličine tela kod izumrlih vrsta
Kao da su sa druge planete
Da smo iz vasione mogli da posmatramo razvoj naše planete i života na njoj, videli bismo da je tokom nekoliko milijardi godina prolazila kroz razne faze. Da smo se nekim slučajem mogli prošetati po njenoj površini, iznenadili bismo se oblicima života koji su se smenjivali. Šetnja po ovakvoj planeti sigurno ne bi dugo trajala jer nije bilo ni kiseonika ni drugih uslova za život. Vremenom je život postajao sve raznovrsniji; među njima je bilo mnogih organizama koji nas neprestano fasciniraju svojim oblikom ili veličinom. Teško je poverovati, ali Zemlja je tokom dužeg perioda svog postojanja prikazivala život kao da je sa druge planete. Razna čudovišta džinovskih razmera su postojala u kopnenim i u dubokomorskim sredinama i vrlo su česti u fosilnim zapisima. Velike dimenzije tela su mogle donositi mnoge prednosti uključujući bolju sposobnost hvatanja plena, uspeh u izbegavanju predatora, inteligenciju, dugovečnost i reproduktivni niz. Veća životinja ima manji odnos površine i zapremine od manjih životinja, što rezultira manjim gubitkom toplote u okolini i omogućava joj da duže opstane u hladnom okruženju. Međutim, jedan veliki nedostatak većih organizama je što su više specijalizovani i potrebno im je više hrane, na primer. Ovo je, između ostalog, dovodilo vrste u veći rizik izumiranja. Veliki broj vrsta, rodova, familija i drugih viših taksonomskih kategorija zauvek je iščezao. Zato se danas ne moramo plašiti; nema dinosaurusa, pterodaktila, a ni džinovskih insekata koji bi nam mogli preletati iznad glave. Ipak, ne možemo a da se ne zapitamo: kako nastaju džinovi i šta oni zapravo predstavljaju ?

Istorija života je složena priča o promenama uslova sredine, načinima prirodne selekcije i faktorima koji određuju koliko prirodna selekcija može ići u nekom pravcu. Nastanak giganata s jedne strane ili patuljastih formi s druge strane, dokazan je brojnim primerima. Džinovi iz geološke prošlosti fasciniraju ne samo običnog čoveka već i naučnike, pa je gigantizam kao česta pojava kod fosilnih organizama razmatrana sa različitih aspekata. Šta se sve svrstava u džinove , divove ili gigante? Životinja može biti džinovska u poređenju sa nama ili džinovska u poređenju sa drugim srodnim životinjama u istoj grupi, pa je prilično zbunjujuće to što se koristi ista reč za označavanje različitih bića. Uspostavljeni su razni kriterijumi prema kojima bi divovi trebalo da budu retki u poređenju sa drugim srodnim životinjama u istoj grupi i „mnogo“ veći od prosečne veličine srodnih vrsta. Na kraju, divovi bi trebalo da budu tipični predstavnici svoje vrste i ne bi trebalo da uključuju atipično velike jedinke uzrokovane patološkim promenama.
Pojave životinja sa velikim telom su proučavane najčešće pojedinačno, ali opšti uslovi koji su omogućili evoluciju i održavanje gigantizma ostali su prilično nejasni. Različiti pristupi ovoj temi dali su različite rezultate. Veličina tela se može izraziti ili kao linearna dimenzija (ukupna dužina ili visina, prečnik) ili kao telesna masa. Definicija gigantizma kao veoma velike telesne veličine je ekološki važna osobina povezana sa različitim faktorima, ali ostalo je pitanje kako ovi faktori mogu da objasne uočeni istorijski obrazac razvoja globalnog gigantizma na nivou ere i na nivou fanerozoika? U literaturi se navode brojni dokazi da je veličina tela primarna posledica ključnih osobina životne istorije kao što su plodnost, mortalitet, stope rasta, metabolizam, način na koji organizmi komuniciraju sa okolinom, dužina vegetacije, kvalitet i količina hrane, kiseonik, klima i dr.
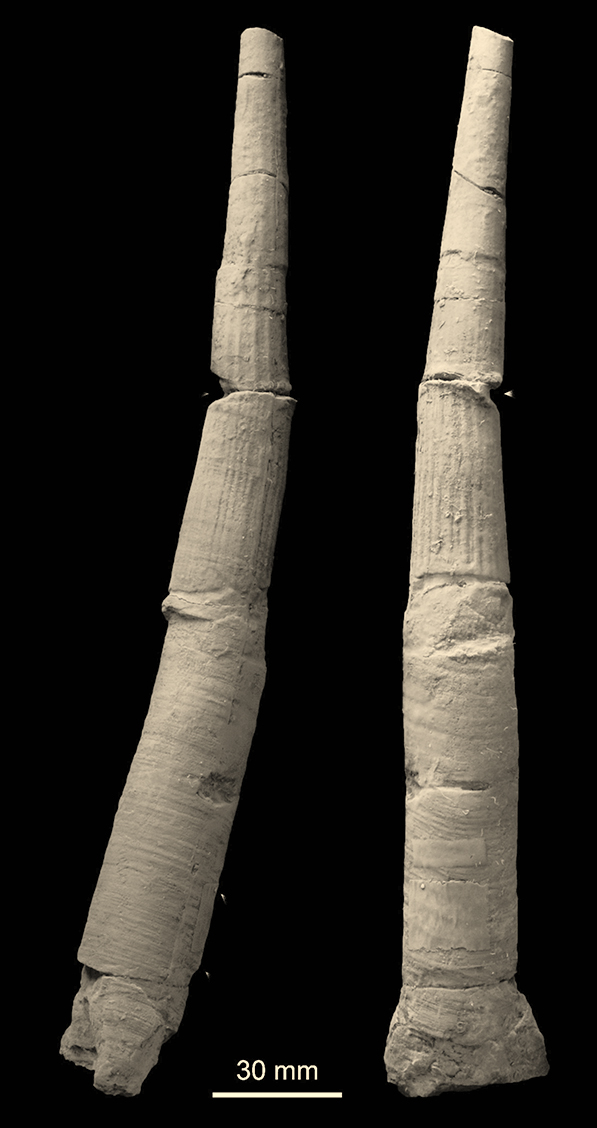
Gigantski skafopod iz perma
|
Na tragu dzinovskih prapotopskih stvorenja
Veličina tela je ključna determinanta različitih fizioloških, anatomskih i ekoloških parametara istorije života, i kao važna biološka osobina svih organizma je proučavana sa raznih aspekata. Fiziologija podrazumeva proučavanje i tumačenje fizičkih i hemijskih faktora odgovornih za nastanak i razvoj života, sa posebnim akcentom na mehanizme koji regulišu životne procese. Mnoge fiziološke karakteristike su verovatno oblikovane ekstremnim uslovima koji su dovodili do izumiranja. Na primer, masovno izumiranje na kraju perma bilo je očigledno manje ozbiljno za životinje tolerantne na visoku koncentraciju CO2. Prestanak aktivnosti tokom jedne ili više životnih faza verovatno je omogućio vrstama da opstanu u vremenima produženog stresa. Ova sposobnost je možda pomogla malim sisarima da prebrode izumiranje krajem krede. Kao posledica brojnih istraživanja i tumačenje uticaja faktora na razvoj velikih tela, predloženo je više hipoteza da objasne mehanizme koji su najbitniji za fenomen gigantizma (npr. Bergmanovo pravilo, Kopeovo pravilo, ostrvsko pravilo itd.).
U fosilnom svetu ima veliki broj primera gigantizma tokom fanerozoika - vremenskog perioda koji obuhvata poslednjih 542 miliona godina, od kada su počele da se pojavljuju prve životinje sa čvrstim delovima. Jedan od najranijih primera gigantizma desio se u periodu brze diverzifikacije života zvanom „kambrijska eksplozija“, kada se pojavila grupa zglavkara poznatih kao anomalokarididi. Smatra se da je Anomalocaris bio grabežljivac i da je mogao narasti do 2 m. U morskom carstvu, procenjuje se da je rani devonski euripterid Jaekelopterus rhenaniae narastao više od 3 m. Euripteride (Chelicerata), poznati kao morski škorpioni iz silura i devona (pre oko 400 miliona godina) su bili veliki vodeni predatori. Imali su izuzetno velike i složene oči. Fosili džinovskih vodenih škorpiona obično se nalaze u bočatnim i slatkovodnim naslagama, mada se pretpostavlja da su verovatno prvo živeli u plitkim priobalnim područjima i estuarima, a kasnije su se preselili u slatkovodne sredine. Najveći kopneni člankonožac, miriapod Arthropleura sličan stonogi, postojao je u periodu karbona (pre 326 miliona godina), dostigao je više od 2 m u dužinu, 55 cm u širinu i bio je oko 50 kg težak. Morao je imati hranu sa visokim sadržajem hranljivih materija, najverovatnije biljnim detritsom u šumskim staništima blizu obale, a možda su bili i grabežljivci koji su se hranili drugim beskičmenjacima, pa čak i malim kičmenjacima kao što su vodozemci. Uzrok njihovog izumiranja je neizvestan, ali bi mogao biti posledica globalnog zagrevanja koje je učinilo klimu previše suvom da bi preživeli, ili možda zbog porasta gmizavaca koji su ih nadmašili u hrani i ubrzo zavladali istim staništima.
Uticaj kiseonika
Kod mnogih istraživača preovlađuje mišljenje da je kiseonik uzročno povezan sa velikim dimenzijama tela. Povećanje kiseonika u atmosferi tokom fanerozoika je povezano sa nekoliko evolucionih inovacija, uključujući prve višećelijske životinje, prve leteće insekte itd. Povišen sadržaja kiseonika u karbonu se, prema nekim autorima, odrazio kod nekih artropoda, kao što je insekt Meganeuropsis, sličan vilinskom konjicu, koji je imao raspon krila od oko 70 cm. Neke stonoge su bile duge 200 cm. Prisustvo gigantizma kod vodenih člankonožaca uključuje i trilobite i rane devonske euripteride (morski škorpioni).
Proučavanja gigantizma u zavisnosti od kiseonika daju različite rezultate u zavisnosti od grupe organizama koja se proučavaju, kao i od toga da li organizmi koriste kiseonik iz vazduha ili iz vode. Mnogi ektotermi u polarnim morima danas su veći od njihovih rođaka sa toplijom vodom. I ako je paleozoik poznat po organizmima sa velikim telom (paleozojski gigantizam) iz vremena sa povišenim koncentracijama kiseonika u praistorijskoj atmosferi, polarni gigantizam (hladni uslovi favorizuju velike veličine tela) se zasniva se na primerima gigantskih ektotermnih organizama pronađenih u ledenim vodama sa velikim količinama rastvorenog kiseonika. Međutim, biološka raspoloživost kiseonika je niža u hladnim vodama, uprkos većoj rastvorljivosti kiseonika. Stoga se primeri polarnog gigantizma ne mogu se uzeti kao podrška paleozojskom gigantizmu.
Pravila, temperatura tela, rast i nestajanje
Bergmanovo pravilo definiše odnose između veličine organizma i temperature u njegovoj životnoj sredini. Prema ovom pravilu, pripadnici jedne vrste ili nekog drugog taksona koji žive u hladnijim oblastima imaju veće dimenzije tela od pripadnika istog taksona koji naseljavaju toplije oblasti. Porast temperature u okolini je takođe predložen kao mogući spoljni razlog za gigantizam nekih izumrlih kičmenjaka, uključujući i zmije (npr. piton Titanoboa i gušteri). Pored toga, temperatura je mogla odigrati veliku ulogu u gigantizmu ostalih kičmenjaka koji su živeli u tropima tokom perioda staklenika, kao što su neki krokodilomorfi iz krede ili miocena. Do nedavno se smatralo da se Bergmanovo pravilo odnosi isključivo za homeotermne organizme. Novija istraživanja ukazuju na prisustvo ovakvih pravilnih promena veličine tela i kod poikilotermnih organizama, ali i na pojavu tzv. inverznog Bergmanovog pravila - smanjenja tela organizma sa smanjenjem temperature sredine.
Jedan od najčešće proučavanih makroevolucionih koncepata je poznat kao Kopeovo pravilo. Generalno, ovo pravilo opisuje povećanje veličine tela tokom vremena. Teorija se obično pripisuje radovima Edvarda Drinkera Koupa u kasnom 19. veku, raspravlja se o tome kako ga treba definisati i koje klade pokazuju primere. Kladistika (od grčke reči κλαδος, klados - grana) je biološka disciplina koja se bavi određivanjem odnosa između organizama na osnovu izvedenih sličnosti. Godine 1973, američki paleontolog Stenli je pokušao da definiše Koupovo pravilo, praveći razliku između onoga što je nazvao aktivnim i pasivnim trendovima. Sa aktivnim trendom, populacija kao celina vremenom raste, a manji pojedinci nestaju; ovo se ponekad naziva Kopovo pravilo u užem smislu. U pasivnom sistemu, srednja veličina se povećava kroz proces u kome maksimalna veličina raste, minimalna veličina ili ostaje konstantna ili se smanjuje, tako da se ukupan raspon mogućih veličina vremenom širi. Ovo se naziva Kopeovo pravilo u najširem smislu. Može se desiti i obrnuto od oba ova obrasca. Koncept je detaljno opisan u mnogim udžbenicima, postoje mnogi predlozi o prednostima velikih tela, ali nijedan do nedavno još nije bio empirijski potvrđen. Koristeći opsežnu analizu mnogih proučavanja, neki naučnici su pokušali da pokažu kako veličina doprinosi preživljavanju, uspehu parenja i plodnosti i iznose argumente za mehanizam koji je sposoban da dokaže Kopeovo pravilo.
Nekoliko različitih faktora olakšava sticanje gigantskih veličina kod predatorskih životinja. Fenomen predator je usko povezan sa fiziološkim ili ekološkim promenama koje su omogućile životinjama da se nose sa povećanim trošenjem energije tokom aktivnog predatorstva. Međutim, zajednički mehanizmi koji leže u osnovi ovog fenomena nisu detaljnije razmatrani. Evolucija anatomskih struktura i fizioloških mehanizama koji omogućavaju stvaranje i/ili zadržavanje toplote, može se smatrati bitnim faktorom koji podstiče razvoj većih tela. Na primer, najveće žive ribe sa aktivnim predatorskim načinom života su mezoterme. Ovaj faktor bi mogao biti uključen u gigantizam nekih izumrlih kičmenjaka predatora kod kojih je nedavno potvrđena endotermija (fiziološka pojava da organizam - endoterm održava telesnu temperaturu uglavnom upotrebom toplote koja se oslobađa tokom njegovih unutrašnjih telesnih funkcija) ili mezotermija (mezoterme - vrste sa umerenim toplotnim zahtevima), uključujući dinosauruse, plesiosauruse. Velika tela i aktivni načini života koji se pretpostavljaju za značajan broj izumrlih vodenih kičmenjaka sugerišu da je pojava endotermije ili mezotermije mogla biti češća u prošlosti nego što se očekivalo.

Gigantoproductus iz karbona
|
Divovi i među beskičmenjacima: stvar ishrane?
Gigantizam fosilnih morskih beskičmenjaka je zapažen kod nekoliko vrsta mekušaca, cefalopoda, brahiopoda, velikog broja krinoida itd. Među mekušcima, skafopodi su prilično retki i imaju vrlo male ljušturice, pretežno nekoliko cm dužine. Paleozojski skafopodi su među najslabije poznatim mekušcima zbog njihove cevaste morfologije ljušture i fragmentarnog očuvanja. Nedavno je ipak pronađeno nekoliko vrsta u permskim krečnjacima Akasaka, u Japanu, koji u odnosu na druge skafopode dostižu velike dimenzije, i više od 20 cm dužine. Među najkrupnijim je poznat skafopod Prodentalium onoi sa velikom ljušturom, dužom od 30 cm. Pretpostavlja se da su postojale velike količine hrane tokom taloženja krečnjaka Akasaka, a samo oni koji su uspeli da je iskoriste su dostigli veliku telesnu veličinu. Skafopodi su mesojedi, hrane se foraminiferima, pa se zaključuje da su se ovi neobično veliki skafopodi hranili krupnim foraminiferima (Fusulinide).
Brahiopodi su se pojavli još u paleozoiku ali, za razliku od mnogih fosilnih grupa poput trilobita i amonita koji su izumrli, raznovrsnost brahiopoda je vremenom opadala. Na neki način nas fasciniraju jer su uspeli da prežive do danas. Ima oko 350 savremenih vrsta i pretežno su malih dimenzija (nekoliko cm). Vrste brahiopoda Gigantoproductus su divovi unutar paleozojskog sesilnog bentosa. Gigantoproductus giganteus je dostizao dimenzije do 30 cm a ostali mogu stati na jedan dlan. Bili su najveći brahiopodi koji su ikada živeli, u svetu brahiopoda bili su pravi divovi. Ovo predstavlja dilemu jer živi brahiopodi imaju niskoenergetski način života. Iako su stope metabolizma brahiopoda verovatno bile veće tokom paleozoika nego danas, ogromna veličina koju su dostigle vrste Gigantoproductus je ipak neobična. Ispitujući ishranu vrsta Gigantoproductus iz karbona Misisipija, zaključeno je da je gigantska veličina koju je dosegao Gigantoproductus verovatno rezultat miksotrofnog načina života, pomoću kojeg su se mogli osloniti na energiju i hranljive materije dobijene iz fotosimbiotskih mikroba i iz filtrirane hrane u obliku čestica.
Da je problematika gigantizma vrlo kompleksna pokazuju interesantna tumačenja istraživača koji su pokušali da otkriju kako je amonit Parapuzosia seppenradensis mogao da naraste do prečnika 1,5-1,8 m. Prethodna istraživanja su pokazala da prosečan amonit nije bio veći od pola metra u prečniku. Istraživači nisu uspeli da pronađu nijedan ubedljiv dokaz koji bi objasnio zašto su počeli da rastu, napominjući da je to moglo biti zbog promene klime. Ali otkrili su da su mnoge vrste reptila mozazaura počele da rastu otprilike u isto vreme. Ovi veliki morski gmizavci su bili glavni predatori amonita. Istraživači sugerišu da je Parapuzosia seppenradensis počeo da raste zbog toga što je mozazaurima bilo teže da ih pojedu, pa su oni veći preživeli i razmnožavali se. Istraživači su ipak na kraju priznali da postoji jedan nedostatak u njihovoj teoriji jer su druga istraživanja pokazala da su i mozazaurusi nastavili da postaju sve veći od kako je Parapuzocia seppenradensis dostigao svoju najveću veličinu. Nakon nekog vremena, ova vrsta je ponovo počela da se smanjuje iz nepoznatih razloga.
Najveći i najstariji pingvin
Ostaci ogromnog, najvećeg do sada pronađenog fosilnog pingvina Kumimanu biceae nađeni su na Novom Zelandu, u stenama kasnog paleocena (pre oko 59 do 56 miliona godina). Kosti krila i nogu ovog giganta su pokazali da se radi o najstarijoj fosilnoj vrsti pingvina visine 1,7 m i težine čak 100 kg. Filogenetski, vrsta Kumimanu biceae predstavlja prilično arhaičnu vrstu, koja se jasno razlikuje od drugih žinovskih pingvina iz geološki mlađih epoha eocena i oligocena. Gigantizam nije bio redak među ranim pingvinima, čak ni u najranijoj fazi njihove evolucije, a zaključuje se da se razvio kao rezultat neletenja morskih ptica. Pošto su se džinovski pingvini razvili ubrzo nakon masovnog izumiranja pred kraj perioda krede, nestanak velikih morskih gmizavaca omogućio je pingvinima da se rašire u nove ekološke niše. Međutim, sa pojavom drugih velikih morskih predatora kao što su foke i kitovi, pingvini su se suočili sa novom konkurencijom i novim predatorima. Ovo je možda dovelo do njihovog izumiranja zbog čega danas nemamo živih divova pingvina.

Najveći pingvin
|
Ostrva su u prošlosti kao i danas stvarala evolucione neobičnosti ili čudne ekosisteme. Ovo takozvano „ pravilo ostrva “ daje opšte objašnjenje za evolucioni razvoj na ostrvima, ali neka pitanja ostaju i dalje sporna. Promene telesne mase sisara uglavnom su objašnjene veličinom ostrva i prostornom izolovanošću, a modulisane klimom, što je rezultiralo izraženijim gigantizmom ili patuljastim formama na malim i udaljenim ostrvima. Dobar primer ostrva i uslova za razvoj giganata sa jedne strane i patuljastih formi sa druge strane je ostrvo Hateg, u Transilvaniji (Rumunija) koje se pre 70 miliona godina nalazilo u moru Tetis. Ekosistem džinovskih pterodaktila koji jedu male dinosauruse čudan je čak i paleontolozima.
Vreme velikih biljojeda
Najvažnija od svih selektivnih prednosti biljojeda je ta što veće dimenzije tela smanjuju pritisak predatora. Većina dinosaurusa među kojima su najveći biljojedi sauropodi bili su veći od najvećeg predatora, teropoda. Njihovo veliko telo je otežavalo neprijatelju da napravi dovoljno snažne i efikasne ugrize.Sauropodi su polagali veliki broj jaja, a mladi sauropodi su bili vrlo male životinje, što znači da su veoma brzo rasli. Sauropodomorfi su bile najveće kopnene životinje zabeležene u istoriji života. Prvi sauropodomorfi su bili mali dvonošci i dugo se verovalo da se sticanje džinovske veličine tela u ovoj kladi (preko 10 t) dogodilo tokom jure i bilo je povezano sa brojnim modifikacijama skeleta koje postoje kod Eusauropoda (pravih sauropoda). Međutim, zbog nepotpunih fosilnih zapisa još nisu proučene ove ključne evolucione promene. Novija razmatranja ističu veoma ubrzanu stopu rasta, poboljšani respiratorni sistem u ptičjem stilu i modifikacije muskulature pršljenova i zadnjih udova kao ključne za evoluciju gigantizma. Ovo otkriva da se prvi impuls ka gigantizmu kod dinosaurusa dogodio više od 30 miliona godina pre pojave prvih Eusauropoda.
"
U poređenju sa drugim biljojedima, dugačak vrat je sauropodima omogućavao efikasnije uzimanje hrane nego kod drugih velikih biljojeda, što znači da su bili u stanju da uzmu više energije iz svog okruženja nego drugi biljojedi. Proučavanjem klime i fosilnih nalaza iz mezozoika, za period od pre oko 230 do 66 miliona godina, zaključeno je da su fosili drugih glavnih tipova dinosaurusa bili prisutni svuda, a mnogi su živeli i u polarnim regionima. Veliki sauropodi dinosaurusi kao što su Brontosaurus i Diplodocus radije su živeli u toplijim, tropskim regionima Zemlje i verovatno su imali drugačiju fiziologiju od drugih dinosaurusa. Staništa sauropoda su verovatno bila otvoreni, polusušni predeli, slični današnjim savanama. Najveći predstavnici svih ostalih dinosaurusa su retko prelazili 10 t, što znači da bili u rasponu veličina veoma velikih kopnenih sisara koji su nastali u kenozoiku, kao što su fosilni indrikoteri i današnji i fosilni slonovi. Među današnjim životinjama, samo kitovi mogu da narastu do telesne mase veće od sauropoda, ali s obzirom da pripadaju različitim sredinama (vodenoj i kopnenoj) nisu pogodni za poređenje.
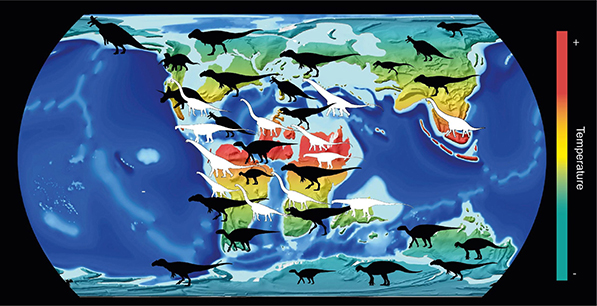
Rasprostraanjenje sauropoda
|
Veličina tela - stvar konkurencije?
Drugi pristup proučavanju evolucije gigantizma odnosio se na proučavanje džinova iz raznih kategorija - najvećih vrsta u svojoj ekološkoj kategoriji ili kladi kroz fanerozoik, razmatranje povlačenja globalnog gigantizma kopnenih životinja u kenozoiku i pojavu maksimalne veličine većine morskih kategorija koje su dostigle svoj vrhunac tokom kasnog kenozoika. Pokušao se dati i odgovor na pitanje: zašto su mezozojski kopneni kičmenjaci postigli veće maksimalne veličine od kasnijih kenozojskih? Analizirane su neke nedoumice kod donošenja zaključaka objavljenih za pojedinačne grupe životinja i proučavani faktori koji su odlikovali vreme posle paleozoika (posebno drugu polovinu mezozojske i kenozojske ere), kada su sve ekološke kategorije bile svedoci evolucije svojih najvećih predstavnika. Analizirano je deset kopnenih i morskih tipova prema mestu ishrane i staništa, ispitani drugi faktori koji opisuju okolnosti koje su bile potrebne za nastanak veoma velikih tela: klima, kiseonik i veličina staništa... što su faktori koji omogućavaju, ali ne primoravaju evoluciju veoma velikih dimenzija tela. Da bi se objasnio gigantizam, najpre se analiziralo rasprostranjenje giganata, a zatim su razmatrani faktori koji su učinili gigantizam mogućim. Analizirani su obrasci gigantizma tokom vremena za nekoliko velikih klasa, uključujući klase mekušaca, nekoliko grupa člankonožaca i nekoliko vrsta kičmenjaka. Pošlo se od toga da je selekcija u korist ekstremnog gigantizma nastala uglavnom zbog konkurencije, a u manjoj meri zbog predatora. Konkurencija je mogla razvijati ekološke međuzavisnosti u lancu ishrane, stvarajućí uslove koji su omogućili da se posle paleozoika razviju neke vrste konkurentno superiornijih životinja u cilju postizanja izuzetno velikih veličina tela. Prema tome, jedna vrsta mora biti najveći član svoje klade, trofičkog nivoa (mesta u lancu ishrane) ili ekosistema, bilo u određeno vreme ili za sva vremena. Lokalni gigant je definisan kao vrsta sa najvećim telom na određenom mestu ili vremenskom intervalu. Vrste koje su najveći članovi svoje ekološke (trofičke i stanišne) kategorije na globalnoj prostornoj skali i/ili u čitavom fanerozoiku nazvani su globalnim gigantima. Analize su vršene za maksimalne veličine tela unutar ekoloških, vremenskih ili filogenetskih kategorija; nisu korišćeni statistički podaci jer statistika opisuje svojstva i razlike između distribucija populacija u odnosu na jednu ili više varijabli kao što je veličina tela.
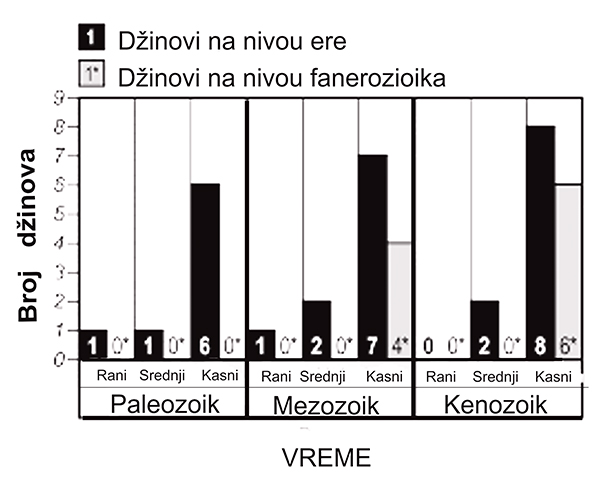
Broj divova na nivou ere i na nivou fanerozika
|
I za žive i za fosilne gigante postoji značajna nesigurnost pri merenju veličine jer maksimalna veličina varira unutar i između lokacija, između mužjaka i ženki, mlade i odrasle forme (što je veoma bitno kad su fosili u pitanju jer ih je ponekad teško razlikovati), a zavisi i od očuvanja fosila i kvaliteta uzorkovanja. Džinovska školjka Crassostrea gryphoides iz nekih lokaliteta evropskih miocenskih mora, na primer, dostiže dimenzije oko 80 cm u dužinu. Ova vrsta je pronađena širom Srbije, uglavnom manjih dimenzija. Na osnovu pronađenih delova ljuštura iz okoline Golupca, mogla se rekonstruisati njihova veličina (oko 0,5 m). Zbog toga su odabirana najveća merenja za najveće vrste u svojoj kategoriji, sa obrazloženjem da je jedinka sa veoma velikim dimenzijama bila u stanju da raste i opstane u barem nekim uslovima u kojima je vrsta postojala. Pokazalo se da su najveće životinje fanerozoika, u svih deset ekoloških kategorija (lanac ishrane i staništa), bile mezozojske ili kenozojske starosti. Svi globalni kopneni giganti na nivou fanerozoika javljaju se od sredine krede do kasne krede; ceo raspon od 100 miliona godina od kasne jure do kraja krede obeležen je ekstremnim gigantizmom kod dinosaurusa (sauropodi, ornidropodi, ceratopsians i hadrosaurus). Iako nekoliko klada na nivou klase dominiraju u redovima giganata na nivou ere (diapsidi u mezozoiku, sisari u kenozoiku i mekušci u obe ere), nema dokaza da se ekstremni gigantizam zadržavao unutar klade u uzastopnim erama. Na primer, paleozojski morski brahiopodi, glavonošci i leteći insekti postali su džinovi na nivou ere, ali ne i tokom kenozoika, uprkos tome što su opstali kao klade na nivou klase. Članstvo u kladi nije važno zbog toga što klada ima svojstva koja pogoduju evoluciji velikih tela, već zato što pojedinačne vrste stiču osobine i žive u okolnostima koje su kompatibilne sa gigantizmom.
Da bi održala veoma veliko telo, životinja mora da ima dovoljno hrane i da do nje lako dolazi. Primarna proizvodnja hrane na morskom dnu je verovatno ostala niska sve do evolucije morskih trava u kasnoj kredi i velikih smeđih algi umerene zone u kasnom paleogenu. Svakako nijedna velika životinja biljojed se nije pojavila u moru pre kasne jure. Stoga bi trebalo da postoji neki opšti vid odnosa između maksimalne veličine tela i produktivnosti životne sredine, brzine kojom hrana postaje dostupna. Biljojedi na grebenima, na primer, značajno su mali u poređenju sa onima koji žive u morskoj travi. Veliki biljojedi koji žive na zemlji ne mogu lako da manevrišu među velikim, gusto raspoređenim drvećem, i ne mogu da dosegnu lišće i grane krošnje. Oni koji žive u drveću ili lete između njih ograničeni su u maksimalnoj veličini u poređenju sa onima koji žive na zemlji.
Dve metaboličke vrste
Dokazi pokazuju da klima igra bitnu ulogu kao faktor koji omogućava ekstremni gigantizam. Visoka rastvorljivost kiseonika i niska potreba za kiseonikom u hladnoj vodi bi trebalo da učine polarne regione i duboko more (i duboka hladna jezera) idealnim za uspostavljanje globalnih divova. Ogromni ranoordovicijumski anomalokaridi i trilobiti iz Maroka i Portugala, koji su u to vreme bili smešteni na višim južnim geografskim širinama, objašnjavani su niskim temperaturama u kojima su živeli. Najveće životinje u svim trofičkim i stanišnim kategorijama održavaju visoku telesnu temperaturu bilo tako što proizvode veliku telesnu toplotu ili žive na toplom mestu. Analize gigantizma kod fosilnih i živih sisara, prema nekim radovima, pokazuju da je maksimalna veličina vrsta kopnenih sisara koncentrisana u vremenu tokom toplog srednjeg eocena, nešto hladnijeg oligocena i promenljivo toplog i hladnog pliocena pa do danas. Međutim, zanemarene su razlike u srednjoj globalnoj temperaturi, na primer između eocena i pleistocena, i ukazuju na činjenicu da su tropski uslovi uvek postojali tokom fanerozoika, što dodatno komplikuje donošenje zaključaka. Globalno hlađenje sa pojavom široko rasprostranjene glacijacije u pleistocenu uticalo je na tropske krajeve, ali mnogi današnji divovi kao što su preživeli slonovi, nosorozi, zmije, kornjače, gušteri, krokodili, insekti, pauci, rakovi, rakovi pustinjaci, stomatopodi, potkovičasti rakovi, nedavno izumrli kopneni lenjivci i izumrli južnoamerički džinovski glodari su rasprostranjeni u tropima. U svakom slučaju, najveći eocenski sisari su manji od kasnijih iz kasnog oligocena i pleistocena.
Uticaj veličine
Što se tiče brzine metabolizma i zastupljenosti kiseonika, mogu se razlikovati dve metaboličke kategorije veoma velikih životinja: životinje sa niskom metaboličkim potrebama, sporim navikama, sporim rastom i često dugim životnim vekom (u prvoj kategoriji su lokalni divovi kao što su džinovske lignje i druge antarktičke životinje koje zauzimaju hladne ili neproduktivne sredine gde su opasnosti od metabolički aktivnih vrsta niske ili povremene) i aktivne životinje sa visokim metaboličkim stopama i energetskim zahtevima, termičkom kontrolom i brzim rastom. Svi najveći pripadnici trofičkih i stanišnih kategorija tokom fanerozoika pripadaju drugoj grupi vrlo aktivnih životinja. Ove osobine omogućavaju velikim životinjama da brzo pređu velike udaljenosti u potrazi za hranom. Veliki kopneni i morski mezozojski gmizavci verovatno su bili mezotermi koji su održavali visoku telesnu temperaturu zahvaljujući svojim velikim dimenzijama tela i čiji se metabolizam približio endotermnom. Velike aktivne životinje zahtevaju obilje kiseonika zajedno sa drugim efikasnim sredstvima za njegovu distribuciju po celom telu. To je dovelo do hipoteze da je gigantizam kod kasnopaleozojskih zglavkara na kopnu bio moguć zahvaljujući nivoima kiseonika u atmosferi koji su bili 20% do možda 33% viši od današnjih. Međutim, uloga nivoa kiseonika u postpaleozoiku ostaje kontroverzna.
Uticaj veličine staništa nije očigledan za sve grupe organizama. Veoma velike pokretne životinje sa velikim apetitima zahtevaju prostrane terene i široku geografsku distribuciju na nivou vrsta. Kenozojske endotermne (toplokrvne) životinje su tako dostigle maksimalne veličine na najvećim kopnenim masama. Megakontinenti mezozoika mogli su da omoguće gigantizam kod dinosaurusa i pterosaurusa; međutim, superkontinent Pangea (od kasnog paleozoika do ranog trijasa) nije omogućio razvoj kopnenih životinja, ni blizu tako velikih kao što su bili kasniji mezozojski ili kenozojski kičmenjaci. Uslovi u unutrašnjosti Pangeje su verovatno bili previše suvi i previše nepovoljni da bi mogle nastati veoma velike kopnene životinje.
S obzirom da veliko telo često daje prednosti u konkurenciji i odbrani od predatora, navodi se da su predatori i konkurenti primarni agensi uticali na razvoj velikih tela kod nekih taksona. Zapaženo je da najveći kopneni biljojedi u većini slučajeva dostižu veće maksimalne veličine od njihovih predatora. Petostruka ili veća razlika između maksimalne veličine biljojeda i predatora, međutim, ukazuje da je malo verovatno da će predatorstvo biti jedini, ili čak najvažniji uzrok koji favorizuje ekstremni gigantizam. Međutim, dominacija u konkurenciji za hranu, sklonište ili parenje kod životinja, za svetlost i vodu, često se povezuje sa telima velikih dimenzija i, za razliku od predatora, ne smanjuje se čak ni kod onih koji imaju velika tela. Predatorstvo je stoga možda bilo važno u ranoj fazi evolucije povećanja veličine vrsta koje su na kraju postale gigantske u eri ili na nivou fanerozoika, ali je konkurencija verovatno učinila da ove vrste dostignu njihove maksimalne veličine.
Džinovska ajkula iz Višnjice
Tokom geološke prošlosti su postojali brojni faktori koji su uticali na pravce prirodne selekcije. Kako se gigantizam razvijao kod nekih grupa organizama, zavisio je od fizioloških i anatomskih inovacija koje su bile najpre korisne kod malih životinja u odbrani i ishrani. Postoje razlike između kopna i mora koje se manifestuju mnogo čvršćom vezom između primarne produktivnosti i gigantizma u morskim ekosistemima nego u kopnenim. Izgleda da je gigantizam kao sredstvo za postizanje konkurentske superiornosti izgubio na značaju sa pojavom grupnog lova, jer grupa relativno malih predatora čini ranjivim čak i veoma veliki plen. Ovaj oblik kolektivnog predatorstva postao je relativno raširen među sisarima mesožderima tokom kasnog kenozoika. Jedinke sa velikim telima imaju prednost u konkurenciji među odraslima, ali životinje veličine mezozojskih džinovskih sauropoda i drugih gmizavaca mogle bi postati lake mete za vrlo pokretene kenozojske endotermne predatorske sisare, posebno za one koji zajednički love.
Organizmi i njihove osobine su proizvodi interakcija koje se menjaju tokom vremena, kako se menjaju uslovi u njihovom okruženju. Pošto je evidentno da se paleozoik po veličini tela razlikuje od mezozoika i kenozoika, sasvim realno je da objašnjenje leži u razlikama u stopi, pristupu i sudbini primarne proizvodnje hrane za koju se oni kojima je neophodna takmiče. Neki faktori mogu da objasne manje maksimalne veličine kopnenih životinja iz kasnog paleozoika i kenozoika, ali neki argumenti dovode u sumnju ovo objašnjenje. Prvo, neki morski sisari su izuzetno veliki uprkos njihovom neefikasnom disanju i kratkim vratovima, kako su ranije bila data tumačenja za neke grupe životinja. Zapažena je smena krupnih predatora u morskoj sredini kroz vreme, kao što je na primer džinovska ajkula poznata pod imenom megalodon (oko 18 m dužine) iz miocena zamenjena sa predatorom kitom ubicom u pliocenu. Džinovska ajkula Otodus megalodon iz Višnjice, kod Beograda, sudeći na osnovu veličine zuba, izgleda da je bila najveća ajkupa u južnom delu Panonskog mora. Veliki je broj analitičkih publikacija često sa kontradiktornim ili nepotpunim zaključcima. Svake godine se na listu fosila dodaje još nekoliko predaka današnjih organizama koji bi vremenom trebali dati bolja objašnjenja o razvoju džinova iz prošlosti naše planete.
Kako su sisari postali dominantna grupa?
Za evolucioni uspeh sisara novi fosili će tek otkriti zamršene grane njihovog evolucionog stabla. Iako se velika otkrića pojavljuju širom sveta, najveći broj dolazi iz Kine; sve je jasnije da je zastarelo verovanje o malim sisarima iz mezozika koji su živeli u senci ogromnih dinosaurusa. Mezozoik je poznat po razvoju dinosaurusa, ali malo je poznato da su se sisari razvijali istovremeno sa njima. Obično se veruje da su to bile male životinje veličine rovki, poput 205 miliona godina starog Morganucodon-a iz Velsa i Kine. Sisari se mogu smatrati velikom grupom (mammaliformes) koja se pojavila u kasnom trijasu, kada su dinosaurusi tek počeli da se diverzifikuju (pre oko 220 miliona godina). Ali mammaliaformes je velika grupa koja uključuje vrste koje su danas potpuno izumrle. Mezozojski prasisari su se javili u mnogim oblicima. Castorocauda je, na primer, bio jurski ekvivalent dabra; to je najranija pronađena životinja sa krznom. Volatikoterijum, približno iz istog vremena, podsećao je na leteću vevericu. Bilo je i onih veličine jazavca kao što je Repenomamus koji su jeli bebe dinosaurusa. To znači da se u kenozoiku ipak nije dogodilo čudo, pa su se sisari naglo razvili. Novi fosili otkriveni u Koral Blufsu (Kolorado) pokazuju kako se Zemlja oporavila nakon masovnog izumiranja dinosaurusa i zašto su sisari postali dominantne životinje. Otkriće je bilo revolucionarno jer je znanje o tome kako su se sisari oporavili pre 66 miliona godina nakon što je asteroid ubio dinosauruse i 75 procenata svih vrsta na Zemlji je bilo simbolično pre ovog istraživanja. Sisari koji su preživeli udar asteroida bili su toliko mali (mnogi nisu bili veći od pacova, pretežno su pronađeni samo fragmenti, delovi oklopa kornjače, poneki krokodilski zub, male vilice sisara,) da je ponovno traženje fosila u trošnim steaama bilo gotovo nemoguće. Na sreću, u zbirkama muzeja su sačuvane konkrecije - stene jajastog oblika koje se formiraju oko fosila, pa su naučnici potražili fosile u njima i pronašli više potpuno netaknutih fosila iz vremena nakon kredno-paleogenog izumiranja, uključujući četiri lobanje sisara. Shvatili su da nalazište Koral Blufs predstavlja prvi milion godina nakon izumiranja dinosaurusa, gde su uspeli da pronađu stenu na dnu Blufsa i stenu na vrhu Blufsa.
Pre udara asteroida, najveći sisari su bili veličine rakuna, a dodatni fosili iz Koral Blufsa otkrivaju povećanje veličine tokom vremena. Kada su neptičji dinosaurusi izumrli pre 66 miliona godina, sisari su opstali. Posle udara asteroida, najveći sisari su bili veličine pacova, da bi im se u svetu bez visokih dinosaurusa ukazale nove mogućnosti. Oko 300.000 godina nakon izumiranja, najveći sisari su bili veličine velikih dabrova. U prvih 10 miliona godina nakon događaja masovnog izumiranja, sisari su se povećavali umesto da razvijaju veći mozak, da bi se prilagodili dramatičnim promenama u svetu oko sebe. Smatra se da, u haosu nakon udara astetroida, veliki mozgovi verovatno ne bi bili korisni za opstanak ranih placentnih sisara. Uspon sisara je zahtevao vreme. Tek su prvi predstavnici savremenih grupa sisara kao što su primati i mesožderi počeli da razvijaju veći mozak i složeniji raspon čula i motoričkih veština. Ove adaptacije su poboljšale njihove šanse za opstanak u vreme kada je konkurencija za resurse postajala sve veća. Mnoge grupe današnjih životinja prvi put su se pojavile u eocenu. Ovo uključuje prve pripadnike slonova, slepih miševa, kitova, parnoprstih kopita (artiodaktila) i neparnih perisodaktila. Prvi konj, Hiracotherium se pojavio u eocenu. To je najraniji član porodice kopitara, koja obuhvata sve izumrle konje i savremeni rod Equus. Tek u eocenu, više od 10 miliona godina nakon udara, sisari su postali zaista veliki i evoluirali u niz zveri koje su bile konkurentne dinosaurusima, ali ipak nisu imali mnogo sličnosti sa svojim savremenim srodnicima. Bilo je potrebno oko 10 miliona godina da sisari biljojedi postanu dovoljno veliki da liče na današnje bizone i antilope.
Džinovi u budućnosti planete
Džinovske životinje predstavljaju izazov evolucionistima koji pokušavaju da razumeju evoluciju veličine tela. Već decenijama se razmatraju razni faktori sa ciljem da se ustanove uslovi razvoja džinova u izumrlom i živom svetu i uzroci njihovog izumiranja. Geološki dokazi pokazuju da će neki od uslova koji se predviđaju za budućnost podsećati na okolnosti koje su bile česte u prošlosti Zemlje. Da bi procenili adaptivne opcije, raspoložive za organizme koji bi se razvijali u takvim okolnostima, poučno je ispitati paleofiziologiju, tj. načine na koje se drevni život suočio sa njegovim fizičkim i hemijskim okruženjem. Da bi to učinili, potrebna su proučavanja koja se zasnivaju na temeljnim principima, kvantitativno potvrđeni kod živih vrsta i mogu se prepoznati na fosilnim ostacima. Ostaje mnogo toga da se sazna o fiziologiji drevnih organizama, posebno o morskim. Jasno je da postoje mnogi načini da se postane div, a svaki novi pronalazak naučnike ponovo navodi da razmišljaju o uslovima koji su omogućili njihov razvoj.
Dr Gordana Jovanović
Kompletni tekstove sa slikama i prilozima potražite u magazinu
"PLANETA" - štampano izdanje ili u ON LINE prodaji Elektronskog izdanja
"Novinarnica"
|